
We tend to think of ways to categorize animals either by shape (4 legs? 2 legs? no legs?, thin? Fat? big headed? small headed?) or phylogenetic affinities (reptiles? lesser animals? Snakes? Skinks?). But what about ecology?
The many aspects of a species ecological niche are generally quantified singly, or we refer to some abstract “multidimensional niche”, meaning we give up trying to characterize it before we even started. In a paper published recently in the Journal of Biogeography GARDians, led by Enav Vidan and Jonathan Belmaker, tried to define, and numerate, the main types of lizards out there – as reflected in their ecology.
The many aspects of a species ecological niche are generally quantified singly, or we refer to some abstract “multidimensional niche”, meaning we give up trying to characterize it before we even started. In a paper published recently in the Journal of Biogeography GARDians, led by Enav Vidan and Jonathan Belmaker, tried to define, and numerate, the main types of lizards out there – as reflected in their ecology.
 Komodo dragon (Photo: Marinus Hoogmoed)
Komodo dragon (Photo: Marinus Hoogmoed) We have selected four traits that we felt define much of the fundamental axes of ecological variation seen in lizards: 1. use of space / microhabitat preference, that defines where in the environment a lizard is active (on the ground? In trees or rocks? Under ground? In water?); 2. Activity times: species active in the same place, or even on the same branch or piece of ground, can segregate their use of the environment by dividing the temporal niche. Furthermore, being diurnal or nocturnal (or being cathemeral and enjoy both ‘worlds’) has strong implication on thermal biology and hence on metabolism and rates in which lizards take up resources and exchange them with the environment; 3. Diet: is a species insectivorous/carnivorous, as most lizards are? Or do they predominantly feed on plant matter (these guys seem to even take more leaves and plant parts with lower energy content)? Or do they in fact use both plants and animals (and then probably energy richer plant parts such as berries or sap)? This is directly related to the way a lizard affects its environment and may also influence its position across the sit and wait-active foraging continuum (no use waiting for plants); 4. Body size – while not an ecological trait per se size nonetheless strongly influences a host of ecological processes, from the degrees of metabolism and energy flow, to the types of available foods – and potential predators.
These four traits obviously interact, and some combinations may be more or less common than others: small, diurnal, terrestrial insectivore is after all the first picture to pop to mind when the term ‘lizard’ is introduced (except for the diehard gecko lovers among us, and well, there are a few of us with this infatuation). But are there tiny nocturnal herbivores? Or huge nocturnal lizards? How common is a marine iguana (large, herbivorous, swimming diurnal beast) type lizard – do we remember it simply because it is exotic?
These four traits obviously interact, and some combinations may be more or less common than others: small, diurnal, terrestrial insectivore is after all the first picture to pop to mind when the term ‘lizard’ is introduced (except for the diehard gecko lovers among us, and well, there are a few of us with this infatuation). But are there tiny nocturnal herbivores? Or huge nocturnal lizards? How common is a marine iguana (large, herbivorous, swimming diurnal beast) type lizard – do we remember it simply because it is exotic?
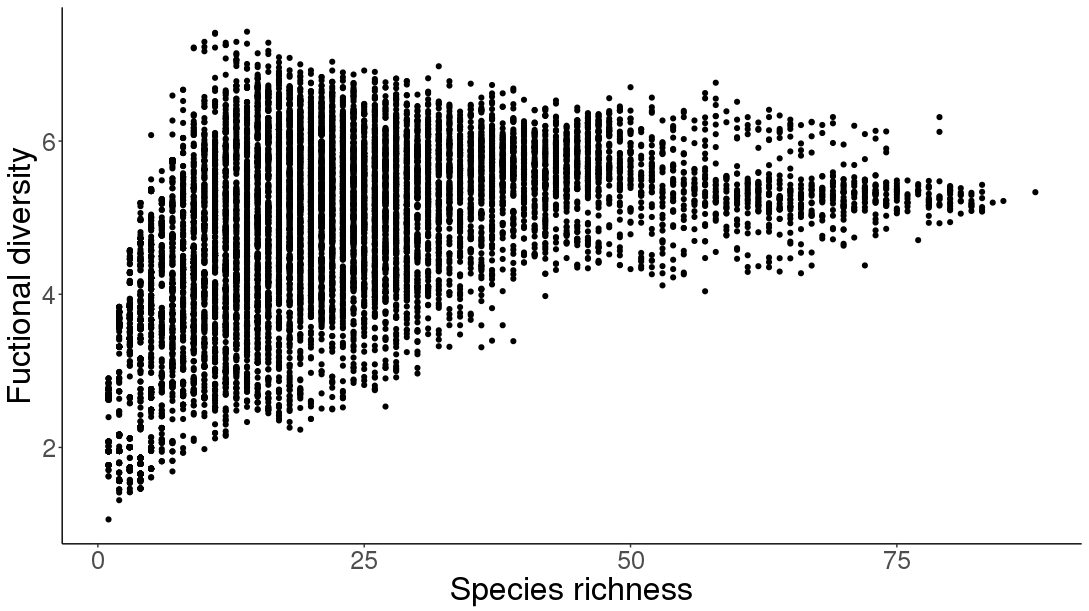
Functional diversity of Lizards as a function of species richness
 Stenodactylus petrii (photo: Marco Ferrante)
Stenodactylus petrii (photo: Marco Ferrante) We used Archetypal Analysis to assign lizards to types – or archetypes. Archetypal Analysis is an unsupervised machine learning technique that seeks to find the number of clusters that create the smallest convex hull in an n-dimensional trait space, by using the extreme values rather than the centroid of the clusters. AA assigns, for each species, a vector of affinities to each archetype. Most species are probabilistically assigned to several archetypes, with the partial probabilities summing to one.
We found that the most common functional trait combinations are (1) diurnal, terrestrial, carnivores (20% of the species); (2) diurnal, scansorial, carnivores (16%); and (3) nocturnal, scansorial, carnivores (13%).
Lizards could be robustly classified into seven ecological “Archetypes”:
We found that the most common functional trait combinations are (1) diurnal, terrestrial, carnivores (20% of the species); (2) diurnal, scansorial, carnivores (16%); and (3) nocturnal, scansorial, carnivores (13%).
Lizards could be robustly classified into seven ecological “Archetypes”:
- terrestrial (obviously, usually small, carnivorous, diurnal thing that are, well, active on the ground), think Ablepharus skinks, for example
- Scansorial – small diurnal, carnivorous, scansorial species, such as Pristurus rupestris.
- Nocturnal – small terrestrial, scansorial and carnivorous species that are, at least partially, active at night (i.e. they are either nocturnal or cathemeral). Like most geckos of course – say Hemidactylus
- Herbivorous - relatively large, diurnal, terrestrial and scansorial species whose diet includes substantial amounts of plant matter (either as omnivores or herbivores). Saharo-Arabian Uromastyx for example fit the bill very nicely
- Fossorial – lizards living at least partially underground, mainly small, carnivorous, with varied activity times. Many skinks, such as Ophiomorus represent this strategy
- Large - very big (all species >200 g), mainly diurnal, terrestrial or scansorial species. You know, Varanus komodoensis and Komodo-dragon wannabes.
- Semi-aquatic - dwelling in aquatic habitats, relatively large, and generally both carnivorous and diurnal, things like Uranoscodon superciliosus
 Uromastyx ornata (photo: Alex Slavenko)
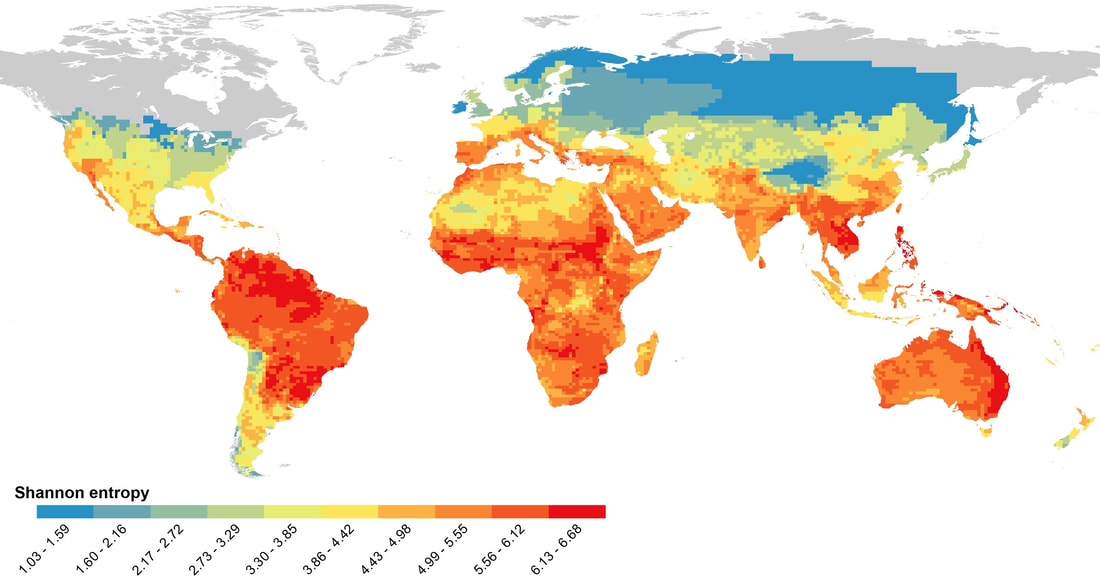
Uromastyx ornata (photo: Alex Slavenko) We then partitioned the global spatial patterns of lizard richness into these seven archetypes, and found that each shows pretty distinct patterns. Turns out Australia is the main hotspot for the herbivorous, nocturnal, fossorial, and terrestrial strategies – and for lizards in general. The Amazon basin is the main hotspot for the semi-aquatic, and scansorial strategies, whereas the large strategy has pan-tropical hotspots, especially in both the Amazon Basin and Northern Australia, but also in Africa, SE Asia and Mexico.
Surprisingly, we found that functional diversity peaks in areas with medium species richness and slowly decreases toward the speciose areas. This unexpected richness-functional diversity unimodal association is also revealed within the scansorial, large, and semi-aquatic strategies. The richness patterns of terrestrial, nocturnal, herbivorous, and fossorial strategies increase with species richness, but globally functional richness peaks in areas with medium species richness.
Thus increases in richness do not necessarily stem from increased functional diversity. Species diversification within specific strategies often dominates richness patterns. Our findings support the contention that it is important to consider different functional and ecological subgroups when studying richness patterns.
Surprisingly, we found that functional diversity peaks in areas with medium species richness and slowly decreases toward the speciose areas. This unexpected richness-functional diversity unimodal association is also revealed within the scansorial, large, and semi-aquatic strategies. The richness patterns of terrestrial, nocturnal, herbivorous, and fossorial strategies increase with species richness, but globally functional richness peaks in areas with medium species richness.
Thus increases in richness do not necessarily stem from increased functional diversity. Species diversification within specific strategies often dominates richness patterns. Our findings support the contention that it is important to consider different functional and ecological subgroups when studying richness patterns.
Author: Shai Meiri
 RSS Feed
RSS Feed
