
 Vallarta Mud Turtle (Kinosternon vogti), classified as ‘Critically Endangered’ by the automated assessment method and as not evaluated by the IUCN Red List of Threatened Species (Photo: Agencia Informativa Conacyt/ Wikimedia)
Vallarta Mud Turtle (Kinosternon vogti), classified as ‘Critically Endangered’ by the automated assessment method and as not evaluated by the IUCN Red List of Threatened Species (Photo: Agencia Informativa Conacyt/ Wikimedia) Reptiles comprise nearly 11,800 species worldwide, and are the most species-rich land-vertebrate group. After 18 years of laborious work by many experts globally, early in 2022, the first extinction risk assessment of this group was completed (the Global Reptile Assessment). This important endeavor will enable adding reptiles to global conservation policy and management initiatives as one of the major groups assessed. Nevertheless, this assessment still leaves over 3000 reptile species that have either not been assessed or were assigned a data deficient category that prevents their prioritization for conservation. In an effort to fill-in this gap a new publication in the journal PLOS Biology, we presents estimates of extinction risk for those species currently neglected by the Global Reptile Assessment, using novel machine learning modelling. importantly we found that unassessed and data deficient reptile species are more likely to threatened than assessed species.
Gabriel Caetano, lead author of the paper explained “The IUCN threat assessment procedure is highly important, yet very lengthy, data intensive, subject to human decision biases, and relies on in-person meetings of experts. However, we can use information on already assessed species to better understand the risks to those not yet assessed. Species may share physiological, geographic, and ecological attributes (often via shared evolutionary history) that make them more threatened, and experience similar sources of threat when they occur at similar locations. In our work we tried to emulate the IUCN process using predominantly remotely sensed data and advanced machine learning methods. We used species that have been assessed to teach our models what makes a species threatened and then predict the threat categories of unassessed species”. He added “our new methods are important for highlighting reptile species at risk and can be used on other groups as an initial shortcut for threat categorization”.
Shai Meiri added “Importantly, the additional reptile species identified as threatened by our models are not distributed randomly across the globe or the reptilian evolutionary tree. Our added information highlights that there are more reptile species in peril – especially in Australia, Madagascar, and the Amazon basin – all of which have a high diversity of reptiles and should be targeted for extra conservation effort. Moreover, species rich groups, such as geckos and elapids (cobras, mambas, coral snakes, and others), are probably more threatened than the Global Reptile Assessment currently highlights, these groups should also be the focus of more conservation attention”
Uri Roll mentioned “Our work could be very important in helping the global efforts to prioritize the conservation of species at risk – for example using the IUCN red-list mechanism. Our world is facing a biodiversity crisis, and severe man-made changes to ecosystems and species, yet funds allocated for conservation are very limited. Consequently, it is key that we use these limited funds where they could provide the greatest benefits. Advanced tools- such as those we have employed here, together with accumulating data, could greatly cut the time and cost needed to assess extinction risk, and thus pave the way for more informed conservation decision making”.
Gabriel Caetano, lead author of the paper explained “The IUCN threat assessment procedure is highly important, yet very lengthy, data intensive, subject to human decision biases, and relies on in-person meetings of experts. However, we can use information on already assessed species to better understand the risks to those not yet assessed. Species may share physiological, geographic, and ecological attributes (often via shared evolutionary history) that make them more threatened, and experience similar sources of threat when they occur at similar locations. In our work we tried to emulate the IUCN process using predominantly remotely sensed data and advanced machine learning methods. We used species that have been assessed to teach our models what makes a species threatened and then predict the threat categories of unassessed species”. He added “our new methods are important for highlighting reptile species at risk and can be used on other groups as an initial shortcut for threat categorization”.
Shai Meiri added “Importantly, the additional reptile species identified as threatened by our models are not distributed randomly across the globe or the reptilian evolutionary tree. Our added information highlights that there are more reptile species in peril – especially in Australia, Madagascar, and the Amazon basin – all of which have a high diversity of reptiles and should be targeted for extra conservation effort. Moreover, species rich groups, such as geckos and elapids (cobras, mambas, coral snakes, and others), are probably more threatened than the Global Reptile Assessment currently highlights, these groups should also be the focus of more conservation attention”
Uri Roll mentioned “Our work could be very important in helping the global efforts to prioritize the conservation of species at risk – for example using the IUCN red-list mechanism. Our world is facing a biodiversity crisis, and severe man-made changes to ecosystems and species, yet funds allocated for conservation are very limited. Consequently, it is key that we use these limited funds where they could provide the greatest benefits. Advanced tools- such as those we have employed here, together with accumulating data, could greatly cut the time and cost needed to assess extinction risk, and thus pave the way for more informed conservation decision making”.
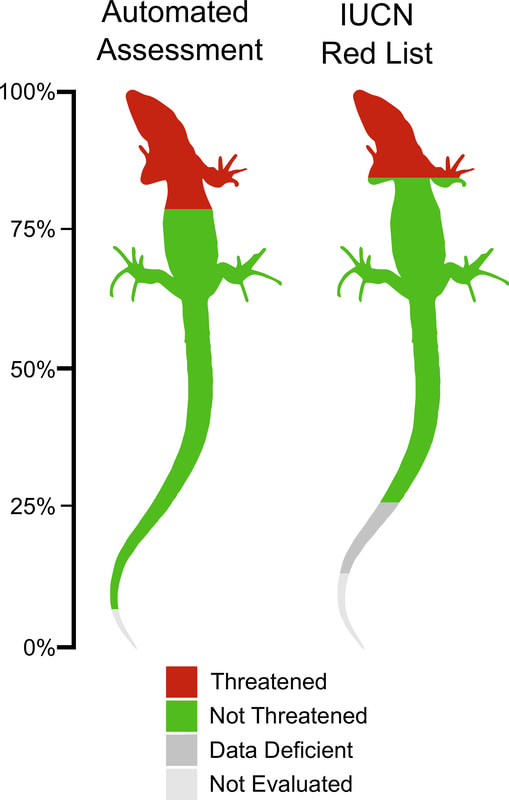
Proportion of reptile species in different threat categories for an Automated Assessment Method and for the IUCN Red List of Threatened Species.







 RSS Feed
RSS Feed
