
In a recent publication in Global Ecology and Biogeography GARDians explored the global diversity and distribution of lizard clutch sizes.

We tested the geographic factors that affect clutch sizes in across nearly 4000 lizard species. We found similar patterns to those that have long been known in birds but were never seriously studied in other groups of organisms: lizards lay large clutches at high latitudes and at highly seasonal regions. We postulate that high latitudes with their short, pronounced productivity peals both allow the production of large clutches and promote putting all the eggs in one basket – because the window of opportunity is short in highly seasonal regions. We hypothesize that this may further be a factor preventing taxa with fixed clutch sizes from colonizing high latitudes.
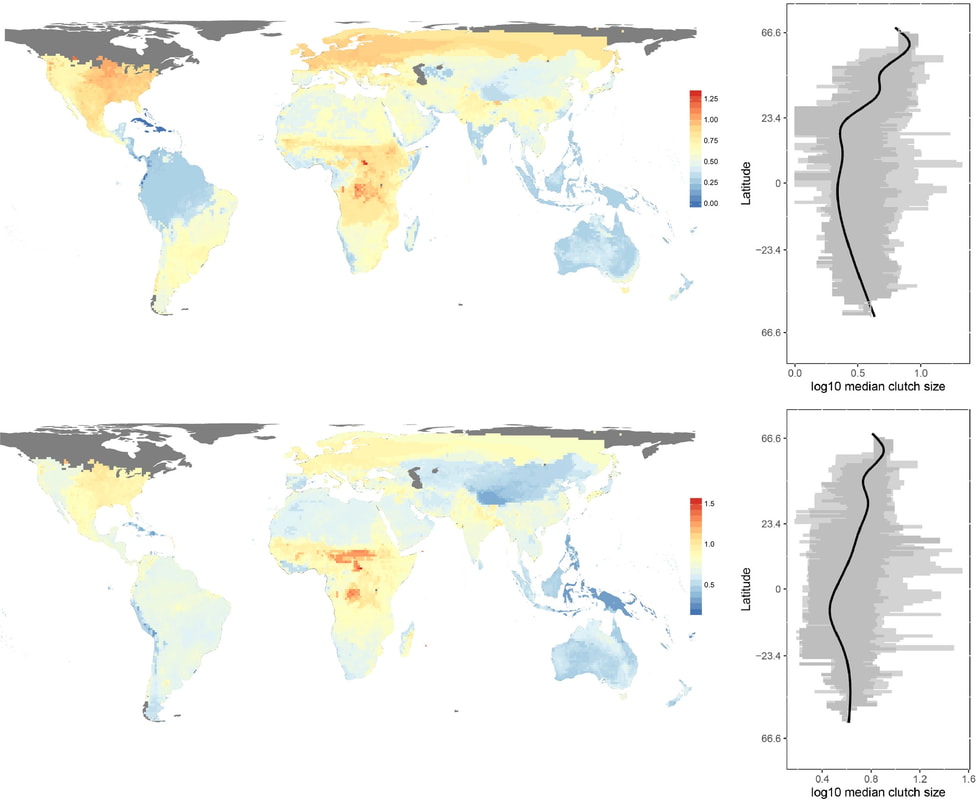
Median log‐transformed clutch size in 96 km × 96 km grid cells globally. Top: all lizards; bottom: only lizards with variable clutch sizes. Note that the colour scale differs between the maps. To the right of each map is a curve showing a generalized additive model of the mapped variable (in black), the 95% confidence intervals of the mapped variable per 96‐km latitudinal band (shaded dark grey), and the range of values of the mapped variable per 96‐km latitudinal band (shaded light grey).
Authors: Shai Meiri, Uri Roll


















 RSS Feed
RSS Feed
